The research in our laboratory interrogates fundamental molecular, cellular and developmental biological processes. In particular, we are interested in the roles played by small ribonucleoproteins (RNPs) in a genetic disease, called Spinal Muscular Atrophy. We are also focused on the functions of histone post-translational modifications (PTMs) in the regulation of eukaryotic gene expression, important for understanding disease mechanisms in many different types of cancer.
Epigenetic control of genome structure and function
Cell cycle-regulated gene expression
Developmental genetics
Synopsis
Our research focuses on understanding the molecular mechanisms that regulate DNA replication and cell proliferation during animal development. An orderly process of events called the cell cycle, which in its most familiar form consists of four phases (G1-S-G2-M) controls cell proliferation. The genome is replicated during the “S” or synthesis phase and duplicated chromosomes are segregated to daughter cells during the “M” or mitotic phase when cell division occurs. G1 and G2 are “gap” phases during cells regulate the entry into S phase and M phase, respectively. We study gene expression events that control how cells make the decision to enter S phase and proliferate, or to exit the cell cycle and differentiate. Without such control there would be no coordination between cell proliferation and the development and function of the many different types of tissues that make up an organism. In addition, the breakdown of cell cycle regulation is one of the events that contribute to the generation of cancer.
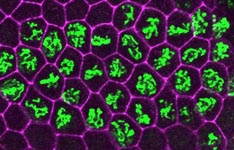
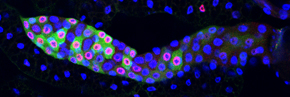
The E2F transcription factor is active (green) during S phase (red) in the asynchronous endocycles of the Drosophila salivary gland.
We study this problem using the fruit fly Drosophila melanogaster, in part because the genes controlling cell proliferation in fruit flies have been highly conserved during evolution and function the same way as in other animals, including humans. This allows us to exploit certain advantages that Drosophila has as a research tool, including the ease with which genetics (making and analyzing mutants) and cell biology (using microscopy to
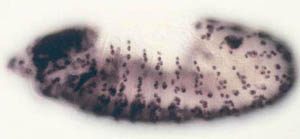
In situ hybridization for histone gene expression in a Drosophila embryo.
observe cell proliferation) can be applied to the study of gene function in the context of a whole animal. Some of the cell cycle regulatory pathways that we study become defective in virtually every human cancer. Thus, one hope is that understanding how these pathways function in normal Drosophila development will give us clues to how they might malfunction in the deregulated growth typical of cancer.
Cellular division, wound healing, chemotaxis, and neuronal outgrowth all rely on dynamic shape change and adaptability afforded via an ever-changing cellular scaffold termed the cytoskeleton. We examine two core components of the cytoskeleton: microtubules and actin filaments in concert with the molecules that regulate them and facilitate communication between them. We employ a combined approach of high resolution time-dependent imaging in parallel with atomic resolution protein crystallography and cryo-electron microscopy to understand, at multiple scales, the molecular processes that control cytoskeletal dynamics. Of particular interest are the +TIP protein families that dynamically localize to growing microtubule plus ends where they regulate microtubule dynamics, communicate with the actin cytoskeleton, capture kinetochores, and engage the cell cortex under polarity-based cues. Investigations proceed through three key areas.
Structure: Tertiary and quaternary molecular architecture of cytoskeletal regulators attained using x-ray crystallography and cryo-electron microscopy.
Cellular and Organismal Imaging: Time-dependent systems analysis via genetic, opto-genetic, and small molecule manipulation.
In Vitro Reconstitution: Microscopy-based physico-chemical analysis of cytoskeletal dynamics and convergent biological events (capture, signaling etc.) through titration of core components and regulators.
Interleaving these efforts, we aim to test, correlate, and bridge information gained from the organismal, cellular, sub-cellular and atomic levels. Of particular interest are the aberrant cytoskeletal molecular mechanisms at play in neuronal disorders and cancer biology.
Ted Salmon is a Cell Biologist and Biophysicist whose primary research is directed towards understanding the molecular mechanisms governing the assembly of spindle microtubules and the segregation of chromosomes during mitosis. Our working hypothesis is that mitosis will be explained by the molecular and structural properties of the centrosome which organizes and nucleates the polymerization of spindle microtubules, the assembly of microtubules which orient and participate in the generation of chromosome movements, and the microtubule motors such as the kinesin and dynein families of proteins which appear to generate polarized forces along the lattice of microtubules, at kinetochores, and within the spindle fibers. The laboratory has pioneered the development of video and digital imaging microscopy methods for analysis of molecular and structural dynamics in living cells and in vitro. We have developed fluorescently labeled tubulins to serve as tracers in studies of the dynamic pathways for microtubule assembly in vivo and in vitro using low light level fluorescence microscopy and digital image processing techniques. We have also developed high resolution video microscopy methods for visualizing the polymerization of individual microtubules and the motility of motors in living cells and reconstituted preparations. These functional assays provide the basis for determination of the basic mechanisms of microtubule polymerization and motor function, analysis of kinetochore motility, and the means of eventually achieving reconstitution of chromosome segregation in reconstituted preparations in vitro. There is also an interest in the lab in how hydrostatic pressure alters the assembly and function of the cytoskeleton and how organisms have adapted to the effects of deep sea pressures.
We are interested in cell-to-cell communication. At present, our efforts are focused on signaling that depends on the heterotrimeric G protein complex. Much of our work is devoted to understanding novel mechanisms for activating this pathway therefore we are looking at organisms quite divergent to animal cells where much is already understood about G protein signaling. Using the diversity of life to discover the diversity of cell signaling is a powerful approach.
The JonesLab showed that most eukaryotic cells outside of the animal group have a very different mechanism for activation mechanism. Whereas, the rate-limiting step for G protein activation is nucleotide exchange on the Galpha subunit of the complex, in plant cells and protists, this step is spontaneous. Therefore, the need for a G protein coupled receptor to catalyze this GTP for GDP nucleotide exchange is not needed in plant cells and protists. We showed that the reaction that returns G protein activation (GTP-bound) to the inactive state is the rate-limiting step and is mediated by a receptor like protein that accelerates this “off” reaction. However, this is not the only mechanism to control the pool size of the activated G protein. The JonesLab takes a wide variety of experimental and theoretical approaches to solve central basic problems in cell biology with the intent that our work will translate into improved quality of life, create sustainable agriculture, and make us better stewards of the Earth. Mathematical modelers, biochemists, microscopists, physiologists, geneticists are welcomed to join the JonesLab team. Please see the available positions by clicking the link above.
(A) G protein regulation in mammalian cells. In the absence of ligand, G protein forms an inactive heterotrimer with GΒγ dimer (left bottom). Ligand-bound GPCR promotes GDP dissociation and GTP binding on G protein (Top). GTP-bound GΒ dissociates from GΒγ dimer, and both activated GΒ and freely-released GΒγ modulate activity of the effectors (right bottom). GΒ hydrolyses GTP to GDP, and re-binds to GΒγ to return to its inactive state. (B) G protein regulation modeled in Arabidopsis. Arabidopsis G protein (AtGPA1) can spontaneously dissociate GDP and activate itself (left bottom). AtGPA1 does not hydrolyze its GDP rapidly, however AtRGS1, a 7TM-RGS protein, promotes the GTP hydrolysis of AtGPA1 (top). D-glucose or other stimuli functions on AtRGS1 directly or indirectly, and decouples AtGPA1 from AtRGS1 (right bottom). Once released from AtRGS1, AtGPA1 does not hydrolyze its GTP efficiently, maintaining its active state, and modulating the effector activities.